Glen Report 25.11: New Product - (5'S)-5',8-Cyclo-dG - DNA Damage and Repair
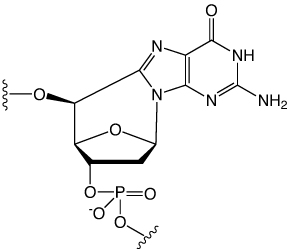
Radiation-induced damage of DNA has been shown to lead to bridged cyclo-nucleosides, with the cyclo-purines, cyclo-dA and cyclo-dG, predominantly formed.
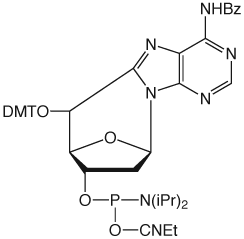
In Glen Report 21.1, Page 10, we introduced 5',8-Cyclo-dA CE Phosphoramidite, (1) on Back Page, in an article that described the repair of DNA damage by Nucleotide Excision Repair (NER) and Base Excision Repair (BER) mechanisms, while focusing specifically on the repair of cyclo-purine lesions. In this article, we update the biological effects of cyclo-purines in DNA while introducing a new Cyclo-dG-CE Phosphoramidite monomer.
The cyclo-purines are formed when a free radical is induced at the C5’ of the deoxyribose sugar by attack by hydroxyl radicals, photolysis or other ionizing processes. The C5’ free radical inserts into the C8-N7 double bond of the adjacent purine residue to form 5’,8-cyclo-purine as either a 5’R or 5’S diastereomer. Of the two diastereomers, the (5’S) cyclo-purine appears to be the most cytotoxic.1 Cyclo-dA is more prevalent than cyclo-dG in DNA damage. Cyclo-purines have been shown to cause significant distortion of the regular DNA helix and these lesions are repaired not by BER but by NER.2,3
In the absence of repair by NER proteins, cyclo-purines accumulate in cells and can block mammalian RNA polymerase II and replicative DNA polymerases.4,5 The distortion in nucleoside structure caused by the C5’-C8 covalent bond perturbs regular hydrogen bonding with the complementary base and has the potential to disrupt normal enzymatic activity in cells. Such DNA lesions may cause polymerase enzymes to be blocked and the efficiency and fidelity of DNA replication may be compromised. The potential for mutations to occur in cells is consequently magnified.6
More recent research on the structure of duplexes containing cyclo-dG has confirmed that base stacking is perturbed at the cyclo-dG site, as well as at adjacent base pairs. Further work7 revealed that mis-insertion of dT or dA opposite the cyclo-dG site can occur. The authors also showed8 that a cyclo-dG…dT mismatch occurs and adopts a wobble base pairing providing a rationale for the observed cyclo-dG -> dA transitions. However, no hydrogen bonding was detected between the cyclo-dG…dA base pair, which is consistent with the observed low levels of cyclo-dG -> dT transitions.
Interestingly, Saccharomyces Cerevisiae and human polymerase eta (pol eta) are able to insert the correct complementary bases opposite cyclo-purines and their adjacent 5' nucleosides at fidelities and efficiencies that are similar to those of their respective undamaged nucleosides. Pol eta’s accurate and efficient bypass of the cyclo-purines contrasts with the mutagenic bypass by other polymerases. The results suggest that pol eta may function in cells to alleviate the cellular burden of endogenously induced DNA lesions, including cyclo-dA and cyclo-dG.9
It is clear that interest in cyclo-dA and cyclo-dG lesions remains high for investigation of cellular DNA damage and repair. We have offered the cyclo-dA monomer since 2009 and we are now happy to add cyclo-dG to our DNA damage and repair toolbox.
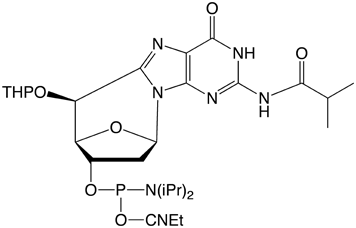
Unfortunately, conventional DMT protection of cyclo-dG is not possible at this time due to severe difficulties encountered in preparing that protected monomer. Instead, 5',8-Cyclo-dG CE Phosphoramidite (2) is offered with a 5’-tetrahydropyran (THP) protecting group. This group is removed by extended treatment with the regular deblock reagent and the subsequent coupling has to be increased to a 15 minute coupling time.
|
|
| Figure 1: Structures of (5'S)-5',8-cyclo-purines | |
References
- I. Kuraoka, et al., J. Biol. Chem., 2001, 276, 49283-49288.
- P. Jaruga, J. Theruvathu, M. Dizdaroglu, and P.J. Brooks, Nucleic Acids Res, 2004, 32, e87.
- P. Jaruga, and M. Dizdaroglu, DNA Repair (Amst), 2008, 7, 1413-25.
- P.J. Brooks, et al., J Biol Chem., 2000, 275, 22355-62.
- B. Yuan, J. Wang, H. Cao, R. Sun, and Y. Wang, Nucleic Acids Res, 2011, 39, 5945-54.
- C. Marietta, and P.J. Brooks, EMBO Rep, 2007, 8, 388-93.
- H. Huang, R.S. Das, A.K. Basu, and M.P. Stone, J Am Chem Soc, 2011, 133, 20357-68.
- H. Huang, R.S. Das, A.K. Basu, and M.P. Stone, Chem Res Toxicol, 2012, 25, 478-90.
- A.L. Swanson, J. Wang, and Y. Wang, Chem Res Toxicol, 2012, 25, 1682-91.
Product Information
8,5'-Cyclo-dA CE Phosphoramidite (10-1098) has been discontinued.
8,5'-Cyclo-dG-CE Phosphoramidite (10-1598) has been discontinued.